Evolution und das Designargument in der Biologie. Ist Intelligent Design in der Natur belegbar?
Gastbeitrag von Martin Neukamm
Die Studiengemeinschaft Wort-und-Wissen vertritt die Auffassung, Lebewesen besäßen Merkmale, die auf einen „geistigen Urheber“ schließen ließen. Natürliche Prozesse wie Evolution seien nicht in der Lage sie hervorzubringen. Widenmeyer/Junker (2016) führen Befunde für diese These an und entfalten ihr Design-Argument (auch „Intelligent Design“ / ID genannt) systematisch. Zudem erklären sie, Kritiker würden am Kern des Design-Arguments scheitern. Nehmen wir die Behauptung ernst und untersuchen, was davon zu halten ist: Wie begründen die Autoren Intelligent Design? Ist ihre Argumentation nachvollziehbar und der Design-Ansatz eine vernünftige Alternative zur wissenschaftlichen Evolutionstheorie?
Als „Design-Ansatz“ bezeichnen die Wort-und-Wissen-Autoren die Vermutung, bestimmte Naturgegenstände (etwa molekulare Maschinen von Zellen) seien ursprünglich auf Schöpfung zurückzuführen. Die Begründung dieser These mithilfe wissenschaftlicher Befunde stellt das Design-Argument dar. Letzteres beruhe im Kern auf der grundlegenden Unterscheidung zwischen „geistigen“ Urhebern und „nicht-geistigen“ Naturprozessen (S. 1).
Nach Widenmeyer/Junker besteht die Begründung für „eine geistige Verursachung und damit für das Design-Argument“ aus zwei Komponenten. Erstens:
„1. Der Naturgegenstand zeigt definierte Kennzeichen von Planung bzw. Zielorientierung (Teleologie), die wir in anderen Fällen ganz entsprechend unseren sonstigen Design-Erfahrungen (Technik, Kunst) ausschließlich auf einen geistigen Urheber zurückführen (wir sprechen dann von ‚Design-Indizien‘ oder ‚Design-Merkmalen‘ wie z. B. funktionale Komplexität; s. u.)“ (S. 1f)
Insbesondere seien „geistig hervorgebrachte“ Gegenstände wie Computer „häufig so gestaltet, dass ihre Teile in z. T. äußerst komplexen und vielschichtigen Zweck-Mittel-Beziehungen stehen“. Entsprechend seien sie oft „hochgradig speziell ausgeführt“, diese Spezifität könne nur „durch solche Zweck-Mittel-Beziehungen erklärt werden“.
„Damit ein Gegenstand eine Funktion ausüben kann, benötigt er in der Regel mehrere Bauelemente, die in spezifischer Weise jeweils konstruiert und zusammen aufeinander abgestimmt sein müssen. Dies betrifft oft ganz verschiedene Aspekte wie Materialeigenschaften, Form, Steuerung u. a. Diese Bestandteile und ihre gegenseitigen Abstimmungen erfordern sehr häufig eine hohe Komplexität, die nicht mehr verkleinert werden kann, ohne die in Rede stehende Funktion vollständig zu verlieren (nichtreduzierbare Komplexität…). Diese Konstruktionen sind an sich klare Design-Kennzeichen…“ (S. 5)
Nichtreduzierbare Komplexität ist ein zentraler Begriff des IDs. Ein System sei nichtreduzierbar komplex, wenn sich keines seiner Bestandteile entfernen lässt, ohne dessen Funktion komplett zu zerstören (Behe 1996, S. 39). Zwar besitzen solche Systeme oft Teile, deren Verlust ihre Aktivität kaum beeinträchtigt. Gleichwohl existiert ein nichtreduzierbar komplexer „Kern“, der für das betreffende System konstitutiv ist.
Lebewesen besitzen zahlreiche solcher Systeme. Sie sollen einerseits ein „positives“ Kennzeichen von Zielorientierung verkörpern. Andererseits seien „die bekannten natürlichen Mechanismen“ „bei weitem überfordert“, sie hervorzubringen. Erklärungen, die auf „überlegte Mittelwahl“ verzichten, ließen „a priori keine Merkmale … erwarten, wie sie bei intentional organisierten Gegenständen vorliegen“ (S. 3). Damit sind wir bei der zweiten (negativen) Komponente des Design-Arguments:
„2. Ein natürlicher Entstehungsvorgang des betrachteten Naturgegenstandes ist unbekannt, und Erklärungsversuche scheitern trotz Wissenszuwachs ...“ (S. 2)
Der Grund, weshalb eine Evolution nichtreduzierbar komplexer Systeme unplausibel sei, besteht Widenmeyer/Junker zufolge darin,
„dass kein kontinuierlicher, kleinschrittiger und hinreichend wahrscheinlicher Weg von einem Zustand ohne diese spezielle Funktion … hin zu einem Zustand mit dieser Funktion (und der dafür nötigen Konstruktion) gedacht werden kann. Jeder einzelne Mutationsschritt müsste eine hinreichende statistische Wahrscheinlichkeit haben. Er dürfte nicht selektionsnegativ sein, vielmehr müsste die entsprechende Mutante in der Population konserviert und signifikant verbreitet werden. Dabei müsste er sich dem in Rede stehenden Zustand annähern … Selbstverständlich kann hier jedoch nicht (schrittweise) auf ein Ziel hin geplant und ‚gearbeitet‘ werden, da natürliche Mechanismen zukunftsblind sind...“ (S. 7)
„Geistige Urheber“ unterliegen dieser Beschränkung nicht, „da sie zielgerichtet vorgehen und vorab im Geiste die verschiedensten Aspekte gleichzeitig beachten und aufeinander abstimmen können“.
„Nach aller unserer Erfahrung gilt: Organisierte Gegenstände entstehen durch den Einsatz von Intelligenz und Planung.“ (S. 5)
Konkrete Design-Argumente, so heißt es, seien naturwissenschaftlich untersuchbar. Ein Design-Argument wäre nach Vorstellung der Wort-und-Wissen-Autoren geschwächt, wenn nachgewiesen würde, dass das betreffende Design-Merkmal natürlich entstehen könne. Umgekehrt stütze ein Scheitern kausaler Evolutionstheorien den Design-Ansatz und weise auf die Grenzen natürlicher Evolution hin (S. 2).
Strukturierter lässt sich das Design-Argument wie folgt darstellen:
These 1: Lebewesen und technische Gegenstände beinhalten (hochgradig) nichtreduzierbar/funktional komplexe Systeme.
These 2: Es ist derzeit kein Fall bekannt, wonach (hochgradig) nichtreduzierbar/funktional komplexe Systeme ohne Design entstanden.
These 3: Ähnliche Eigenschaften haben ähnliche Ursprünge.
Schluss auf die vorläufig beste Erklärung (Abduktion): Die (hochgradig) nichtreduzierbar/funktional komplexen Systeme bei Lebewesen gehen wahrscheinlich auf Design zurück.
Nichtreduzierbare, funktionale Komplexität als Design-Merkmal
Es ist unstrittig: Biologische Systeme sind oft hochgradig nichtreduzierbar komplex. Und nichtreduzierbar/funktional Komplexes erweckt oft den Eindruck, es sei „designt“ worden (Beweis des ersten Anscheins). Doch ein solcher Prima-facie-Beweis ist kein echter empirischer Beleg, sondern zunächst nur ein Verdacht. Um ihn zu erhärten, benötigen wir zusätzliches Erfahrungswissen (Hintergrundwissen), das den Artefakt-Charakter der Objekte stützt (vgl. Mahner 2018).
Nehmen wir an, wir fänden einen faustgroßen, beidseitig behauenen Hornstein mit durchgehend gleichmäßig retuschierten Lateral-Kanten, die eine komplett umlaufende, spitzwinklige Schneide bilden (Abb. 1). Ohne Zögern würden ihn Archäologen als Artefakt einstufen. Das liegt aber nicht allein an der funktional-komplexen Form des Steins.

Der „Design-Charakter“ erschließt sich uns primär aufgrund des Erfahrungswissens, dass Steine mit entsprechender Form und Schneide, passender Größe, aus geeignetem Material usw. traditionell als Werkzeuge dienten und Menschen sie tausendfach herstellten. Nur wenn wir dieses Wissen haben, können wir die (spezifische) Komplexität des Steins als Design-Kennzeichen werten. Dazu gesellt sich die begründete Vermutung, dass die Objekteigenschaften in ihrer Komplexität selten natürlich entstehen; das spielt eine Rolle, wenn wir solche Stücke gehäuft finden. Wäre der Stein hingegen überdimensioniert oder winzig oder zeigte die Schneide Rinde, widerspräche das unserem Erfahrungswissen. Design wäre, trotz funktionaler Komplexität, nicht belegt.
Generell spielt bei technischen Gegenständen wie Computer-Platinen, Botschaften übermittelnden Radiosignalen, Steinköpfen, Uhren und Zahnrädern der Wiedererkennungswert eine wichtige Rolle beim Schluss auf Design. Zudem handelt es sich um Gegenstände, die sich weder selbst zusammenbauen noch reorganisieren.
Bei natürlichen Systemen ist das anders, auch wenn sie nichtreduzierbar komplex sind. Lebewesen unterliegen der Selbstorganisation, Variation und Auslese, sind also evolutionsfähig. Dies schließt nicht aus, dass sie anfangs „designt“ wurden, ja vielleicht designt werden mussten. Nichts aus der Erfahrung spricht jedoch dafür.
Betrachten wir ein Beispiel aus der nichtbelebten Welt: Der globale Wasserkreislauf und dessen Verzahnung mit atmosphärischen und kosmischen Bedingungen ermöglicht Leben auf der Erde. Diese „Feinabstimmung“ lässt sich als eine Form nichtreduzierbarer Komplexität auffassen (Drendel 2016). Trotzdem sehen wir darin nicht die Handschrift eines Designers, denn die Bedingungen auf der Erde sind ein kosmischer Glückstreffer. Sie entstanden über Jahrmilliarden aufgrund von Ereignissen, die nicht abzusehen waren. Der Weg dorthin verlief nicht zielorientiert geradlinig, und auf einen lebensfreundlichen Planeten kommen Tausende unbewohnbarer. Dieser sinnlose Aufwand ist für zielgerichtetes Vorgehen (intelligente Planung) untypisch.
Wir sehen: Die These, ähnliche Eigenschaften (nichtreduzierbare/funktionale Komplexität) hätten ähnliche (intelligente) Ursprünge (causa aequat effectum), ist nicht allgemein gültig. Bei technischen Gegenständen, wo unser Hintergrundwissen für ihre Artefaktnatur spricht, ist Komplexität ein klares Design-Indiz. Bei natürlichen Systemen, die völlig anders organisiert sind und wo dieses Wissen fehlt, ist das nicht der Fall.
Natur- versus Handlungsteleologie (Intentionalität)
Es kommt etwas hinzu: Viele biologische Merkmale lassen sich zwar teleologisch deuten: Das Sekret des Bombardierkäfers erfüllt „den Zweck“, Angreifer abzuwehren. Die Blütenstrukturen von Orchideen „dienen“ dazu, Bestäuber anzulocken. Heliotrope Pflanzen wenden ihre Blütenblätter der Sonne zu, „um“ die Energieproduktion zu steigern. Doch solche Naturteleologie und die sich in den biologischen Merkmalen spiegelnden „Zweck-Mittel-Beziehungen“ unterscheiden sich fundamental von der in Artefakten erkennbaren Handlungsteleologie eines Planers (Toepfer 2004, S. 425).
Eine Handlungsteleologie spiegelt sich in Objekten wider, denen wir die geistige Vorwegnahme eines äußeren Zwecks ansehen. Sie existieren nicht „um ihrer selbst willen“, sondern stehen im Dienst eines Planers. Uhren beispielsweise sind fremddienliche Werkzeuge, von Menschen für Menschen gemacht; ihr äußerer Zweck ist die Zeitmessung. In Kenntnis des Design-Zwecks schließen wir sofort auf einen Zwecksetzer. Bei Lebewesen hingegen ist ein solcher Schluss nicht offensichtlich. Sie weisen nur intern zweckmäßige Strukturen der Selbstorganisation und Reproduktion auf, die „um ihrer selbst willen“ existieren und sich selber hervorbringen. Mangels Handlungsteleologie lässt sich ein Planer nicht ohne weiteres begründen. Es stellt daher
„… eher eine Irreführung als eine gerechtfertigte methodische Parallelisierung dar, wenn beide Bereiche mit der einheitlichen Begrifflichkeit der Teleologie, insbesondere mit dem Wort Zweckmäßigkeit, erschlossen werden sollen.“ (Toepfer 2004, S. 425)

Lässt sich Design ohne zusätzliches Designer-Wissen feststellen?
Der Philosoph Elliot Sober (2008) betont, es sei in der Biologie unmöglich, Design festzustellen, ohne unabhängiges Wissen über mutmaßliche Planer zu haben. Ein unbekannter Zufallsfaktor würde die Herkunft des Auges genauso gut oder schlecht „erklären“ wie ein unbekannter Schöpfer, über dessen Absichten und Handlungsoptionen wir nichts wissen. Widenmeyer/Junker widersprechen dem, indem sie die Mord-Analogie aufgreifen: Wir könnten einen Mord feststellen, ohne den Täter zu kennen; Hilfshypothesen über den Verursacher seien „offenkundig nicht erforderlich“ (S. 14).
Des Weiteren kontern sie Sobers Einwand mit einer Reductio ad absurdum, indem sie das Auge exemplarisch durch einen Computer ersetzen. Sie behaupten, Sober müsse auch für diesen Fall folgern, dass ohne zusätzliches Designer-Wissen nicht entschieden werden könne, ob Computer durch Zufall oder Planung entstünden. Da dies absurd wäre, sei die Forderung nach zusätzlichem, unabhängig testbarem Hintergrundwissen über den Planer und seine Methoden irrelevant.
Wie Mahner (2018, S. 206) bemerkt, gehen solche Vergleiche jedoch am Sachverhalt vorbei: Wir haben genügend unabhängiges Hintergrundwissen über Morde und Mord-Instrumente, „sodass etwas, das nach Mord aussieht, oft auch schnell als Mord klassifiziert werden kann, obwohl wir den Mörder nicht kennen“. Hätten wir dieses Wissen nicht, könnten wir allenfalls konstatieren, dass der Fall wie Mord aussähe, ihn aber ohne zusätzliches Täterwissen nicht als Mord bestimmen. Entsprechend ist auch Widenmeyer/Junkers Computer-Analogie verfehlt. Hier bringen wir ebenfalls so viel Hintergrundwissen mit, dass wir ohne Kenntnis von Herstellern und Technologien auf „Design“ schließen: Wir wissen, dass Computer Menschenwerk sind, welchen äußeren Zwecken sie dienen und dass ihre Schaltkreise weder vom Himmel fallen noch evolvieren.
Auch Außerirdische, die „völlig andersartige Geräte benutzen und von Menschen nichts wissen“ (S.13), hätten die Chance, einen Computer oder Mars-Rover als „gemacht“ zu erkennen. Das setzt nicht primär das Erkennen von „Zweck-Mittel-Relationen“ voraus, sondern die Fähigkeit, ihr Hintergrundwissen über die Unterschiede zwischen künstlichen und natürlichen Systemen auf die Objekte anzuwenden. Sie würden feststellen, dass Mars-Rover lediglich „bewegende Kraft“ haben, keine „in sich bildende Kraft“ (Toepfer 2004, S. 333). Sie würden erkennen, dass sie weder intern zweckmäßige Strukturen besitzen noch evolvieren. Damit wäre Design plausibel.
Design als prüfbare Gegenthese zur natürlichen Entstehung
Widenmeyer/Junker glauben, die Design-These lasse sich mithilfe der Gegenthese überprüfen: Würde im Detail die Möglichkeit eines natürlichen Entstehungsvorgangs naturwissenschaftlich nachgewiesen, der zum „Design-Merkmal“ führt, verlöre das Design-Indiz „seine Kraft“ und „bekäme Konkurrenz“ (S. 2). „Kausale Evolutionstheorien“ würden dies bislang nicht leisten. Dies stütze „das Design-Argument, da es die direkte Gegenthese einer rein natürlichen Entstehung“ sei (S. 9). Doch das stimmt nicht.
Ein Scheitern „kausaler Evolutionstheorien“ würde nicht die Design-These stützen, da mehrere Alternativen in Betracht kämen, wie etwa der Vitalismus des 19. Jahrhunderts (Hemminger 2019). Umgekehrt wäre die Design-These nicht geschwächt, selbst wenn bewiesen wäre, dass höheres Leben durchweg natürlich aus Einzellern evolvierte:
„Intelligentes Design ist durchaus kompatibel mit der Sichtweise, dass das Universum ohne Überschreitung der Naturgesetze funktioniert, wobei das Design des Lebens vielleicht in seine ursprüngliche Struktur gepackt wurde.“ (Behe 2008, S. 166; dt. MN)
Die Annahme, der Planer lasse die Bioevolution ohne Überschreitung von Naturgesetzen ablaufen, ist genauso plausibel oder beliebig wie die Voraussetzung, die Design-These sei die Gegenthese zu „nicht-geistiger“ Evolution. Solange wir über ein Design natürlicher Systeme gar nichts wissen, ist jede dieser Zusatz-Annahmen willkürlich.
Außerhalb der Technik können wir mangels Hintergrundwissen konkrete Design-Thesen also nicht direkt prüfen. In der Natur funktioniert eine Prüfung des Design-Ansatzes nur indirekt über den „Umweg“ einer ausgearbeiteten Theorie: Wir benötigen die explizite Kenntnis potenzieller Designer und ihrer Mechanismen um beurteilen zu können, ob Design vorliegt. Mahner (2018, S. 122):
„Um den Design-Ansatz glaubhaft zu machen, benötigt man neben Hintergrundwissen über die Absichten des Designers genau wie die Evolutionstheorie spezifische mechanismische Erklärungen als konkrete Modelle, die zeigen, welcher Designer auf welche Weise irreduzibel komplexe Organe geplant und ins Leben gerufen hat.“ (Mahner 2018, S. 122)
Diese Forderung zur „weiteren Spezifizierung“ des Design-Ansatzes ist nicht „unbegründet“, wie Widenmeyer/Junker (S. 12) behaupten. Die Gründe sind, dass Design in der Biologie nicht offensichtlich ist und die spezifizierende These, der Designer besitze „ähnliche geistige Merkmale wie Menschen“ (S. 10), nicht reicht, um den Design-Ansatz zu stützen. Eine rein „geistige Verursachung“ gibt es nicht; jeder Designer muss bestimmte Eigenschaften aufweisen, um planungsfähig und handlungsfähig zu sein, sodass wir auf Betrachtungen seiner Natur nicht verzichten können:
„Dann kann man aber auch prüfen, ob dem Verweis auf einen Designer überhaupt eine mögliche Erklärungskraft zukommen kann, d. h., ob ID überhaupt in der Lage ist, ein alternatives Erklärungsparadigma anzubieten.“
Letzteres ist bei natürlichen Systemen nicht der Fall, die positive Komponente des Design-Arguments ist gescheitert. Hintergrundwissen, das Biosysteme als „gemacht“ ausweist, haben wir nicht. Potenzielle Planer kennen wir nicht. Über ihre Techniken, Fähigkeiten und Grenzen wissen wir nichts. Der Design-Ansatz ist spekulativ wie inhaltsleer und keine Alternative zur wissenschaftlichen Evolutionstheorie.
Nichtreduzierbare Komplexität als Einwand gegen Evolution
Kommen wir zur zweiten Komponente des Design-Arguments, der Skepsis gegenüber der natürlichen Entstehung nichtreduzierbar komplexer Biosysteme. Widenmeyer/Junker begründen sie mit vorgeblicher Unwahrscheinlichkeit. Die Grundzüge ihres Arguments stammen von Behe (1996); dieser beschreibt „molekulare Maschinen“, die eine physiologische Funktion erfüllen. Da sich ihr nichtreduzierbar komplexer Kern an keiner Stelle vereinfachen lässt, ohne dass die Funktion wegbricht, sei es unmöglich, ihn in hinreichend kleinen, von der Selektion belohnten Schritten aufzubauen.
In diesem Zusammenhang sprechen ID-Vertreter vom „direkten Darwin‘schen Weg“:
„Ein direkter darwinistischer Weg ist einer, wonach sich ein System durch natürliche Selektion entwickelt, die eine gegebene Funktion schrittweise verstärkt.“ (Dembski 2003; dt. MN)
Eine solche schrittweise Optimierung sei bei nichtreduzierbar komplexen Systemen unmöglich, da sich eine noch nicht vorhandene Funktion nicht verstärken lässt. Zwar ist sich Behe im Klaren darüber, dass der Evolution indirekte Wege offenstehen, um nichtreduzierbar Komplexes zu erzeugen. Eine Möglichkeit ist die Einbettung von vorhandenen Merkmalen bzw. Genen in neue, zusätzliche Funktionszusammenhänge. Doch bildeten sich auf diese Weise nach Ansicht der ID-Vertreter allenfalls sehr einfache Systeme (Kojonen 2016, S. 65f).
Menuge (2004, S. 104f.) begründet dies damit, dass deren Evolution an mehrere Voraussetzungen geknüpft sei: Die Teile, aus denen die neue „Maschine“ entstehe, müssten in der Zelle vorhanden sein und für evolutive Experimente zur Verfügung stehen. Des Weiteren müsse sie die Evolution an den richtigen Ort „setzen“ und passend verschalten. Und sie müsse alle Teile hochgradig speziell feinjustieren, um die Funktion zu gewährleisten. Schon bei einfachen Systemen soll dies vielfache Abstimmungen gleichzeitig erfordern. Die Entstehung relativ komplexer Systeme sei daher unwahrscheinlich.
Erörtern wir das Argument an einem Beispiel: Verschiedene Organellen tierischer und pflanzlicher Zellen (Euzyten) entstanden durch Verschmelzungen ursprünglicher Zellen. Diesen Sachverhalt beschreibt die Endosymbionten-Theorie, wonach eine bakterienähnliche Vorläuferzelle ein Sauerstoff nutzendes Bakterium (Proteobakterium) aufnahm. Ein daraus hervorgehender Eukaryot verleibte sich später ein Photosynthese betreibendes Cyanobakterium ein. In Jahrmillionen büßten die aufgenommenen Bakterien (Symbionten) ihre Unabhängigkeit ein und lagerten den Großteil ihrer Gene in den Kern der Wirtszelle aus. So wurden die Proteobakterien zu Mitochondrien der Tier- und Pflanzenzellen und die Cyanobakterien zu Chloroplasten der Pflanzen (Abb. 2).
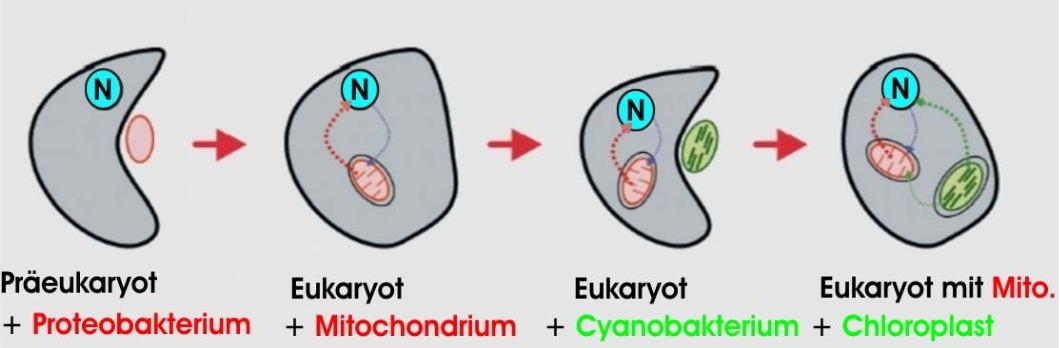
Obwohl die Endosymbionten-Theorie wohlbestätigt ist, hält Wort-und-Wissen (2008) die „Ko-Evolution“ von Symbiont und Wirt für unwahrscheinlich, da ein funktionaler Gentransfer gleichzeitig viele aufeinander abgestimmte Koordinationsschritte erfordere:
- Die Symbionten-Gene müssten sich passend ins Kerngenom der Wirtszelle integrieren, damit sie „lesbar“ sind.
- Die Produkte (Proteine) der in den Kern transferierten Gene müssen wieder in die Symbionten (bzw. Organellen) gelangen. Diesen „Re-Import“ gewährleisten spezielle Transit-Peptide, die den Proteinen angehängt werden müssen. Dafür benötigen die Gene passende Zielsequenzen.
- Es braucht hochspezifische Sortiersysteme (Poren-Komplexe), welche die Proteine in die jeweiligen Organellen „einfädeln“.
- Bevor ein Protein durch den Poren-Komplex gelangt, muss es ein sogenanntes Chaperon entfalten. Anschließend muss eine Protease das Transit-Peptid abschneiden und sich das Protein korrekt zurückfalten (Abb. 3).
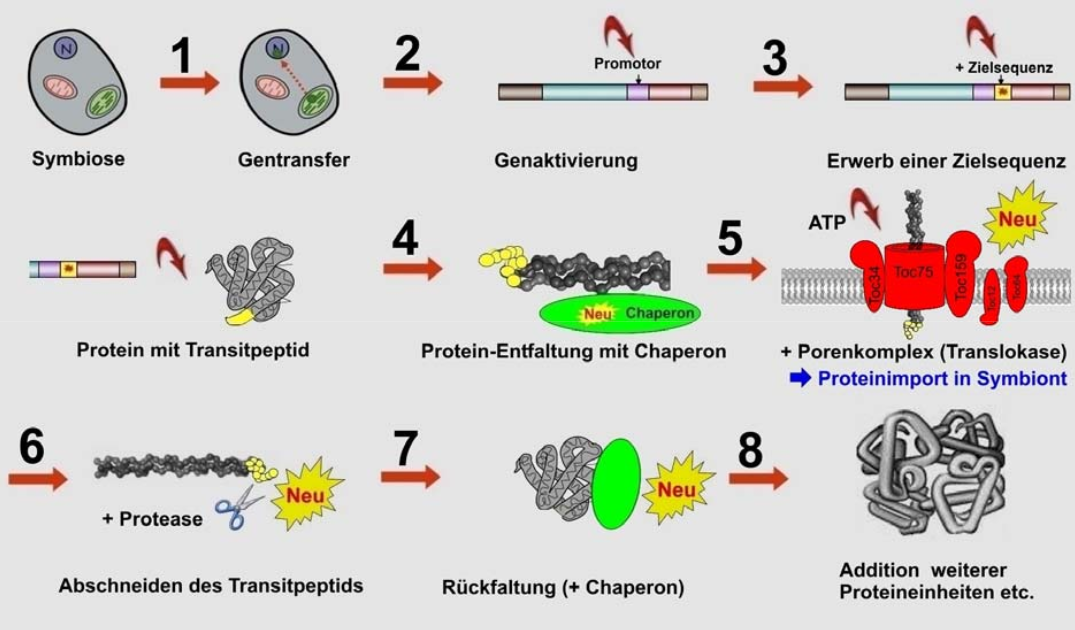
Junker & Scherer (2006, S. 185) resümieren:
„Diese Prozesse müssen alle zusammen gleichzeitig funktionell sein, damit ein kerncodiertes Protein ins Organell transportiert werden und das entsprechende mitochondriale Gen verloren gehen kann. Über den Mechanismus zur Entstehung einer solchen ‚konzertierten Aktion‘ kann derzeit nur spekuliert werden. … Das ganze System erscheint irreduzibel komplex und wirkt als Design-Signal.“
Reduzierbare Spezifität contra nichtreduzierbare Komplexität
Die Pointe an der Sache: Die Forschung konnte zeigen, dass die ersten Endosymbiose-Systeme die spezifischen Einrichtungen heutiger Organellen gar nicht benötigten:
- Zuallererst ist eine komplexe Kooperation zwischen Wirtszelle und Endosymbiont unnötig: Die Vorfahren der Mitochondrien beispielsweise konnten durch ihre bloße Anwesenheit Sauerstoff entgiften.
- Der Gentransfer in den Kern der Wirtszelle kann zunächst unspezifisch erfolgen, wenn er Duplikate betrifft. Dies belegen Hülsenfrüchtler, bei denen Zwischenformen funktionalen Gentransfers existieren (Neukamm & Beyer 2011).
- Experimente zeigen, dass Chloroplasten-Gene überraschend häufig in den Zellkern „springen“. Ein Teil der Pflanzen schafft es, die nichtfunktional in den Zellkern transferierten Gene durch zufälliges Hinzufügen oder Entfernen eines kleinen Stücks DNA zu aktivieren (Stegemann & Bock 2006).
- Zielsequenzen waren für die in den Kern der Wirtszelle ausgelagerten Gene zunächst entbehrlich. Beispiele belegen, dass ein Teil der Gen-Produkte in nicht „vorgesehene“ Zell-Kompartimente gelangen oder im Plasma bleiben kann.
- Die später erworbenen Zielsequenzen brauchten keineswegs besonders spezifisch zu sein: Zwanzig Prozent der kodierenden Gen-Bereiche und ein hoher Prozentsatz von Zufallssequenzen (!) eignen sich dafür (Tonkin et al. 2008).
- Einfache Sortiersysteme sind nicht auf die „enorme Komplexität und Spezifität“ (Wort-und-Wissen 2008) heutiger Poren-Komplexe angewiesen. Zwar sind die Protein-Untereinheiten heute nur in ihrer speziellen physiologischen Vernetzung funktional. Wir wissen jedoch, dass ein Großteil davon ursprünglich unnötig war.
Betrachten wir exemplarisch den Poren-Komplex in der Außenmembran von Chloroplasten (Abb. 4, rechts). Sein Kernstück ist das Protein Toc75. Es steuert gemeinsam mit den Untereinheiten Toc34, Toc159, Toc64 usw. den Re-Import bestimmter, von den Chloroplasten benötigter Proteine.
Beim Cyanobakterium (Abb. 4, links) findet sich das ähnliche Protein SynToc75. Doch es fehlen die Untereinheiten. Es sitzt bereits an der „richtigen“ Stelle und ist in der Lage, bestimmte Proteine passieren zu lassen, wenngleich nicht mit der hohen Spezifität heutiger Chloroplasten. Dies macht SynToc75 zum idealen Ausgangspunkt in der Evolution des Protein-Transporters. Die Evolution konnte dessen Effizienz und Spezifität langsam durch sukzessives Addieren von Rezeptoren und Regulationsproteinen steigern (Reumann et al. 2005; Inaba & Schnell 2008; BodyŁ et al. 2009). Erst später wurde das Gesamt-System für die Zelle unentbehrlich.
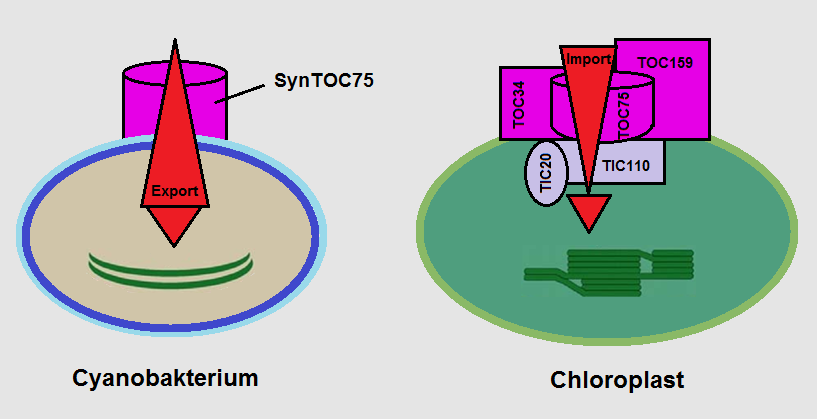
Anders gesagt:Ein hoher Grad an nichtreduzierbarer Komplexität in einem hochspezialisierten System sagt nichts darüber, wie komplex das Arrangement in einem noch wenig spezialisierten „Mini-System“ sein kann. Die Teile der betreffenden Systeme müssen nicht zwingend von Anfang an existieren und „hochgradig speziell ausgeführt“ sein:
„Was Behe braucht, ist nicht nur Spezifität, sondern das, was ich als ‚nichtreduzierbareSpezifität‘ bezeichnen möchte. Die Behauptung, dass die Spezifizität eines Systems nichtreduzierbar sei, entspricht der Behauptung, dass das System aufhört wirksam zu arbeiten, wenn seine Teile modifiziert werden. … Kein System funktioniert so, dass eine Veränderung seiner Teile, egal wie geringfügig sie auch sein mag, zu einem totalen Funktionsverlust führen würde.“ (Draper 2002; dt. MN)
Im Übrigen zeigt sich oft, dass sich viele Bauelemente schon vor der Entstehung der betreffenden Systeme an den „richtigen“ Stellen befanden und prinzipiell funktionsfähig waren. Mehrfach abgestimmte Koordinationsschritte sind nicht zwingend. Nichtreduzierbare Komplexität impliziert daher nicht per se die Unwahrscheinlichkeit von Evolution.
Fairerweise räumt Wort-und-Wissen heute ein, die „schrittweise Entwicklung zu Endosymbionten“ werde „zunehmend plausibel“ (Junker & Scherer 2013, S. 199). Von „Design-Signal“ ist keine Rede mehr. Also ist die Pauschalbehauptung, „die bekannten natürlichen Mechanismen“ seien „bei weitem überfordert“, hochgradig nichtreduzierbar Komplexes hervorzubringen, haltlos. Leider hält das Wort-und-Wissen nicht davon ab, seine unzutreffende Argumentation auf Basis von Wissenslücken (etwa beim Wiesen-Salbei) stetig neu aufzulegen.
Martin Neukamm
Publiziert im März 2020
Für kritisches Gegenlesen und hilfreiche Korrekturvorschläge dankt der Autor herzlich Dr. Martin Mahner von der GWUP sowie Prof. Andreas Beyer von der FH Recklinghausen.
Eine pdf-Version des Artikels bietet die AG EvoBio.

Sie lesen lieber aus einem Buch? Sie finden diesen Artikel auch in unserem dritten Buch zu dieser Webseite, "Gottes Schöpfung und menschliche Technik" (Darmstadt 2022). 17 namhafte Autoren führen den Dialog mit Wissenschaft und Technik angesichts der Gottesfrage weiter.
Literatur
Behe, M. (1996) Darwin's black box. The biochemical challenge to evolution. Free Press, New York.
Behe, M. (2008) The Edge of evolution. The search for the limits of Darwinism. Free Press, New York.
BodyŁ, A. et al. (2009) Early steps in plastid evolution: current ideas and controver-sies. Bioessays 31, S. 1219–1232.
Dembski, W. A. (2003) Evolution's logic of credulity. www.arn.org/docs/dembski/wd_logic_credulity.htm
Draper, P. (2002) Irreducible Complexity and Darwinian Gradualism. A reply to Mi-chael J. Behe. Faith and Philosophy 22, S. 3–21.
Drendel, K. (2016) The irreducible complexity of the universe.
pemiwader.blog/2016/01/28/the-irreducible-complexity-of-the-universe
Hemminger, H. (2019) Warum „intelligentes Design“ keine Naturwissenschaft ist. www.theologie-naturwissenschaften.de/startseite/leitartikelarchiv/intelligent-design
Inaba, T. & Schnell, D. J. (2008) Protein trafficking to plastids: one theme, many va-riations. Biochemical Journal 413, S. 15–28.
Junker, R. & Scherer, S. (2006) Evolution. Ein kritisches Lehrbuch. 6. Auflage, Weyel-Verlag, Gießen.
Junker, R. & Scherer, S. (2013) Evolution. Ein kritisches Lehrbuch. 7. Auflage, Weyel-Verlag, Gießen.
Kojonen, E. R. V. (2016) The Intelligent Design debate and the temptation of scien-tism. Routledge, London.
Mahner, M. (2018) Naturalismus: Die Metaphysik der Wissenschaft. Alibri-Verlag, Aschaffenburg.
Menuge, A. J. L. (2004) Agents under fire: Materialism and the rationality of science. Rowman & Littlefield Publishers, Lanham.
Neukamm, M. & Beyer, A. (2011) Die Endosymbiontentheorie. Allgemeine Grundlagen, Fakten, Kritik.
ag-evolutionsbiologie.net/pdf/2011/Endosymbiontentheorie.pdf
Reumann, S. et al. (2005) Evolution of the general protein import pathway of plastids (review). Molecular Membrane Biology 22, S. 73–86.
Sober, E. (2008) Evidence and evolution: The logic behind the science. Cambridge University Press, Cambridge.
Soll, J. & Schleiff, E. (2004) Protein import into chloroplasts. Nature Reviews Mo-lecular Cell Biology 5, S. 198–208.
Stegemann, S. & Bock, R. (2006) Experimental reconstruction of functional gene transfer from the tobacco plastid genome to the nucleus. The Plant Cell 18, S. 2869–2878.
Toepfer, G. (2004) Zweckbegriff und Organismus. Königshausen & Neumann, Würz-burg.
Tonkin, C. J. et al. (2008) Evolution of malaria parasite plastid targeting sequences. PNAS 105, S. 4781–4785.
Widenmeyer, M. & Junker, R. (2016) Der Kern des Design-Arguments in der Biologie und warum die Kritiker daran scheitern. www.wort-und-wissen.de/artikel/a22/a22.pdf
Wort-und-Wissen (2008) Die Endosymbiontenhypothese. evolutionslehrbuch.info/index2.php
Bildnachweis
Titelbild: Mitochondria. 3d image von Wire_man © Adobe Stock
Abbildung 1: Faustkeile. © Naturhistorisches Museum Toulouse, CC BY-SA 3.0; https://de.wikipedia.org/wiki/Faustkeil#/media/Datei:Biface_Cintegabelle_MHNT_PRE_2009.0.201.1_V2.fond.jpg
Abbildung Auge: close up of human eye von Herr Hesse © Adobe Stock
Abbildung 2: Evolution der Euzyte © Rudolf Bock. Mit freundlicher Genehmigung des Autors
Abbildung 3: Die acht Hauptschritte in der Ko-Evolution © Martin Neukamm
Abbildung 4: Toc- und Tic-Proteine © Martin Neukamm, umgezeichnet nach Soll & Schleiff (2004, S. 204).
Diesen Beitrag fand ich...
Kein Design in der Natur?
Eine Einladung zur Diskussion
In seinem Gastbeitrag macht Martin Neukamm aus naturalistisch-wissenschaftlicher Perspektive deutlich, warum der in manchen kirchlichen Kreisen verbreitete Ansatz von Wort und Wissen nicht trägt. Können Sie seine Argumentation nachvollziehen? Und was halten Sie von der Idee einer geistigen Verursachung, also von Design in der Natur?









Kommentare (3)-

Antworten
Ebenso wie oft kritisiert wird, dass man mit "Evolution" ganz verschiedene Dinge meinen kann trifft das auch auf ID zu und schafft in meinen Augen viel Verwirrung.
-

Antworten
Hallo Frank,
-

Antworten
Ein schöner und fundierter Beitrag. Zwar wird er Kreationisten nicht überzeugen (weil das - faktisch - grundsätzlich unmöglich ist), aber es ist eine gute Zusammenfassung für Interessierte, die auf der Suche nach Argumenten betr. ID sind. Natürlich kann man jedes der Beispiele zerpflücken - weil jedes der Themen so umfangreich ist, dass eine Behandlung in einer solchen Zusammenstellung notwendigerweise didaktische Verkürzungen erzwingt. Bin gespannt, wie lange es dauert, bis das passiert...
Frank Ansari
am 01.03.2020Mir erscheint der Begriff "ID" als "Nachfolgemodell" des Kurzzeitkreationismus (Young Earth Creationism, YEC).
Also etwas so: "OK, wir geben ja nun auch zu, dass die Erde nicht nur 10.000 Jahre alt ist und gemeinsame Abstammung aller Lebewesen ist wohl auch richtig."
Wenn ich Bücher von Michael Behe lese kommt es zumindest bei mir so an (und der dürft in der Szene jan nicht ganz unwichtig sein).
Nun frage ich mal: was ist denn an ID so grundsätzlich anders als das, was man früher "Theistische Evolution" genannt hat?
Die noch spannendere Frage ist aber: was nützt das eigentlich vom theologischen Standpunkt her?
Eines der theologischen Argumente gegen Evolution war doch immer: wenn Evolution war wäre, dann hätte es ja Tod, Krankheit, Leid, Fressen und Gefressenwerden (lauter unschöne Dinge) alle schon vor dem Sündenfall gegeben.
Im Bezugsrahmen von YEC ist das ganz anderes: hier gibt es gar keine "Erdgeschichte" (bzw. diese ist mit der Existenz des Menschen auf diesem Planeten identisch).
Denn offensichtlich hat es in einem erdgeschichtlichen Szenario (und das aktzeptiert ID ja) Tod, Gifte, Raubtiere alles das eben schon seit Urzeiten gegeben.
ID würde also maximal verhindern dass man komplexe biologische Strukturen einem unpersönlichen Prozess zuschreibt (womit bei der Enstehung von Bergen, Flüssen und Seen interessanterweise niemand ein Problem hat).
Auch bei der wissenschaftlichen Aufklärung der Ontogenese wird nicht der Atheismus ausgerufen.
Fazit: ID mag vielleicht ein Lückenbüßer für bisher unverstandene Prozesse sein, lässt aber theologisch so viel verbrannte Erde hinter sich, dass mir bisher unklar ist wie man die Scherben aufräumen soll.
Würde man von einem x-beliebigen Gott ausgehen, wäre das alles gar kein Problem (der könnte ja machen, was er will) - wenn wir aber davon ausgehen (und so ist zumidest mein theologischer Kenntnisstand), dass alles was in der Welt schlecht ist (Tod, Krankeit, Leid, usw.) nicht Teil der ursprünglichen Schöpfung ist dann frage ich mich wie ID da weiterhelfen soll.
Martin Neukamm
am 01.03.2020Du denkst viel zu weit ;-)
All das, was Du ansprichst, ist richtig. Denk aber daran, dass es in diesem Beitrag um das Design-ARGUMENT geht. Also nicht um eine bestimmte Theologie oder Gottesvorstellung, sondern schlicht um "Design-Erkennung". Wenn eine intelligente Ursache in der Natur erkennbar wäre, dann wäre zwar "Design" noch immer unterbestimmt und kausal unverstanden, würde dem Kreationismus wenig bringen usw. Gleichwohl wäre dann ein Design in der Natur plausibel. Religiösen, die den Anspruch vertreten, "Design" empirisch-wissenschaftlich zu stützen, reicht das, denn sie scheinen zu wissen, dass mehr mit wissenschaftlichen Mitteln nicht möglich ist. Ein (konsistentes) Design-Argument ist zwar nicht hinreichend für ein bestimmtes Gottesbild aber notwendig, wenn man das naturalistisch-wissenschaftliche Weltbild überwinden möchte.
Gruß, Martin
Prof. Dr. Andreas Beyer, Vorsitz AG EvoBio.
am 06.03.2020